Ochrilidia gracilis et acrida turrita
III.10.2.6. Ochrilidia gracilis
Les résultats l’étude quantitative de la consommation chez les imagos d’Ochrilidia gracilis sont consignés dans le Tableau 32.
Tableau 32. Recouvrement global, surface moyenne, taux de consommation et indice d’attraction des plantes consommées par 50 individus d’Ochrilidia gracilis en milieu cultivé de Touzouz.
| Espèces | RG (%) | S (mm) | T (%) | IA |
| Cynodon dactylon | 4,91 | 140,5 | 24,84 | 5,06 |
| Setaria verticillata | 3,43 | 117,3 | 20,74 | 6,05 |
| Polypogon monspeliensis | 1,08 | 98,5 | 17,42 | 16,13 |
| Anisantha sterilis | 0,29 | 78,3 | 13,84 | 47,72 |
| Digitaria commutata | 0,20 | 80,5 | 14,23 | 71,15 |
| Hordeum murinum | 0,39 | 50,5 | 8,93 | 22,90 |
| 6 | 10,3 | 565,6 | 100 | 169,01 |
RG : Recouvrement global ; S : surface moyenne ; T : taux de consommation ; IA : indice d’attraction Parmi les six Poaceae consommées par Ochrilidia gracilis, on note un taux de consommation élevé pour Cynodon dactylon (24,8 %) et Setaria verticillata (20,7 %).
Par contre l’indice d’attraction de ces deux graminées est faible (5,1) et (6,05) et leurs taux de recouvrement respective est plus élevé (4,9 %) et (3,4 %).
Digitaria commutata a un taux de recouvrement le plus faible parmi les espèces végétales consommées par ce Gomphocerinae. Cependant, cette graminée marque une valeur importante de (71,15) pour l’indice d’attraction. Le faible taux de consommation est enregistré chez Hordeum murinum (8,9 %) (Figure 28).
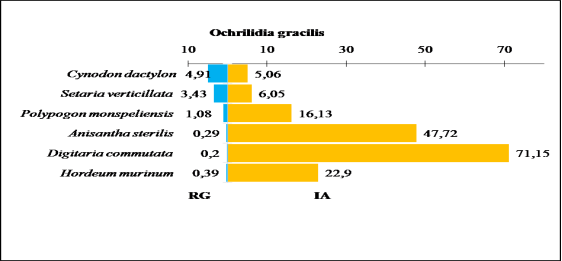
Figure 28. Comparaison entre les indices d’attraction des espèces végétales ingérées par Ochrilidia gracilis et leurs taux de recouvrement au niveau du milieu cultivé Touzouz.
III.10.2.7.Acrida turrita
Les résultats l’étude quantitative de la consommation chez les imagos de d’Acrida turrita sont consignés dans le Tableau 33.
Tableau 33. Recouvrement global, surface moyenne, taux de consommation et indice d’attraction des plantes consommées par 50 individus d’Acrida turrita en milieu cultivé de Touzouz.
| Espèces | RG (%) | S (mm) | T (%) | IA |
| Cynodon dactylon | 4,91 | 192,5 | 28,51 | 5,81 |
| Setaria verticillata | 3,43 | 168,6 | 24,97 | 7,28 |
| Polypogon monspeliensis | 1,08 | 112,5 | 16,66 | 15,43 |
| Anisantha sterilis | 0,29 | 106,3 | 15,75 | 54,31 |
| Phragmites australis | 0,78 | 95,2 | 14,1 | 18,08 |
| 5 | 10,49 | 675,1 | 100 | 100,91 |
RG : Recouvrement global ; S : surface moyenne ; T : taux de consommation ; IA : indice d’attraction
Le taux de consommation relativement important pour les cinq Poaceae, mais avec une certaine tendance pour Cynodon dactylon (28,5 %) et Setaria verticillata (25 %).
Par contre leurs indices d’attraction sont respectivement les plus faibles (5,8) et (7,3). L’indice d’attraction le plus important est enregistré chez Anisantha sterilis (54,3). Le taux de recouvrement de cette graminée par contre est le plus faible (0,3 %) (Figure 29).
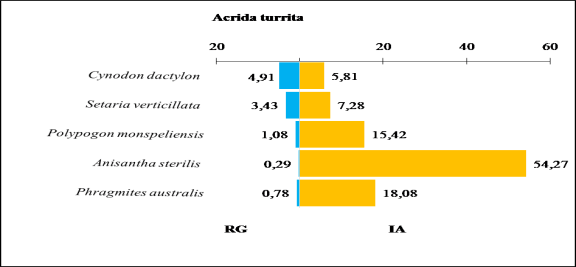
Figure 29. Comparaison entre les indices d’attraction des espèces végétales ingérées par Acrida turrita et leurs taux de recouvrement au niveau du milieu cultivé Touzouz.
Les résultats obtenus par l’étude quantitative montrent qu’Acrotylus patruelis, Aiolopus strepens, Morphacris fasciata, Ochrilidia gracilis et Acrida turrita présentent un taux de consommation élevé pour Cynodon dactylon. Par contre, l’indice d’attraction de cette Poaceae est très faible pour les cinq Acridiens.
Cependant les plantes les plus ingérées ne sont pas forcément les plus appétissantes. Nous avons montré aussi, que la plus part des Acridiens ont consommées au moins une Poaceae, ce qui peut être expliqué selon Singer & Stireman (2001) par l’abondance de ces végétaux dans la nature.
Ces résultats semblent en accord avec ceux de Essakhi et al., (2015). De plus les graminées possèdent moins de composés secondaires (pyrrolizidines, quinolizidines et tannins) que les dicotylédones, qui sont généralement répulsifs pour les acridiens (Capinera et al., 1997).
D’autre part, le bon équilibre en eau des plantes est un facteur essentiel de leurs utilisations. Certes, un apport localisé d’eau d’irrigation, inscrit dans un contexte aride joue également un rôle important dans le maintient de ces Poaceae en turgescence (Ould Elhadj, 2001).
Nos résultats montrent aussi, qu’une espèce végétale moins abondante dans le biotope peut être très attirée par les criquets. C’est le cas d’Anisantha sterilis chez Acrida turrita, Morphacris fasciata et Acrotylus patruelis ; de même pour Digitaria commutata chez Ochrilidia gracilis.
D’après Launois-Luong (1975), le choix de nourriture chez les criquets n’est pas lié à l’abondance des plantes ingérées. Le choix de la nourriture d’acridiens peut être influencé par d’autres facteurs, tels que molécules de dissuasion ou de stimulation (Le Gall, 1989), architecture des plantes et couleur (Picaud et al. 2002).
Certaines de ces fonctionnalités peuvent être détectées par des organes sensoriels, tels que les organes olfactifs ou gustatifs des sensilles des antennes (Chen et al., 2003), chimiorécepteurs sur les palpes (Jin et al., 2006), et des sensilles du labre (El Ghadraoui et al., 2002 ; Zaim et al., 2013).
La taille du corps des criquets devrait être liée à la quantité de biomasse consommée afin de soutenir les demandes métaboliques de ces insectes (Schmitz & Price, 2011).
En effet les grands Acridiens tels que Acrida turrita et Heteracris ont ingérés une quantité de nourriture plus importante que les moins grands tels que Pyrgomorpha cognata et Acrotylus patruelis.
La biomasse consommée est quantifiée en somme moyenne des surfaces ingérées. Les résultats relatives à la quantification de l’alimentation et 1′ analyse de l’indice d’appétence confirment la forte sélectivité des sept espèces Acridiennes à l’égard des plantes disponibles dans leur biotope.
De plus cette étude quantitative a donnée à chaque plante consommée son importance réelle. C’est dire qu’une plante peut être fréquente dans les fèces d’un criquet, mais à des quantités faibles. Parmi les sept Orthoptères analysés, cinq sautériaux s’attaquent uniquement aux plantes adventices.
Par contre Heteracris annulosa et Pyrgomorpha cognata consomment des plantes cultivées telles que Lagenaria siceraria, Cucurbita maxima, Cucumis melo, Solanum melongena, Solanum lycopersicum, Capsicum annuum et Mentha pulegium.
Les dégâts sur ces plantes sont importants au fur et à mesure que la densité d’Heteracris annulosa et Pyrgomorpha cognata augmente dans les parcelles de ces cultures.
De plus cet Eyprepocnemidinae et ce Pyrgomorphinae sont phytophiles. Cependant, Acrotylus patruelis, et Morphacris fasciata sont rarement observés sur végétation, ils sont géophiles.
En outre, les spécialistes d’un habitat risquent l’extinction locale lorsque leurs plantes hôtes et / ou leurs habitats sont modifiés ou détruits car ils ne s’adaptent pas facilement aux changements de plantes hôtes ou d’habitats (Keller et al., 2013).
En effet Heteracris annulosa et Pyrgomorpha cognata sont très liées à leurs plantes hôtes. La densité de ces criquets diminue très considérablement avec le changement des cultures.
Conclusion
A l’issue de ces trois années (automne 2016 jusqu’au fin été 2019) de suivis, pas moins de 31944 d’individus d’Orthoptères ont été comptabilisés sur les 9 stations des 3 sites de la vallée du M’Zab appartenant à l’étage bioclimatique saharien à hiver doux.
Ces spécimens sont représentés par 47 espèces, 28 genres, 15 sous familles, six familles et deux sous ordres. Il a fallu pour leur collecte 324 sorties de prospections et 3240 quadrats de 25 m2. Ce travail nous a permis d’ajouter onze espèces nouvelles pour la faune
Orthoptérologique de la vallée du M’Zab. Ils s’agissent de Notopleura saharica, Heteracris littoralis, Heteracris minuta, Aiolopus puissanti, Aiolopus simulatrix, Hilethera aeolopoides, Morphacris fasciata, Oedaleus senegalensis, Sphingoderus carinatus, Sphingonotus paradoxus et Tropidopola cylindrica.
La sous famille des Oedipodinae est la plus représentée avec 40,4 % des espèces inventoriées. Le genre Sphingonotus est le plus riche avec sept espèces, soit 14,9 % des Orthoptères recensés.
Les méthodes non paramétriques d’estimation de la richesse en espèces réelle ont indiqué que l’inventaire de chaque habitat était complet. Au sein de neuf biotopes inventoriés, les milieux cultivés composent les habitats les plus riches et les plus peuplés en Orthoptères.
En effet, la présence d’une strate herbacée telle que Cynodon dactylon et Setaria verticillata favorise le développement des criquets. Au contraire, la présence d’une trop forte proportion d’Arbres le cas des palmeraies, contribue à la diminution de la faune Acridienne.
Nos résultats montrent que la richesse des espèces atteint un maximum en Eté. Elle est plus élevée dans les sites moins perturbés. En effet ces résultats confirment la réaction des Acridiens aux modifications de la structure de la végétation et du sol suite à l’intensité agricole dans un biotope donné.
Les densités maximales de population d’Orthoptères caractérisent les relevés effectués en milieux cultivés en saison estivale (163 individus à 118 individus pour 100 m²).
Inversement, les densités les plus faibles (3 à 4 individus pour 100 m²) correspondent aux milieux naturels et les palmeraies durant l’hiver. Les densités étaient plus élevées dans les sites moins perturbés de Ben Isguen et Ghardaïa.
Les données sur l’abondance relative des espèces ont montrées que, Morphacris fasciata, Ochrilidia gracilis, Acrotylus patruelis, Aiolopus strepens et Pyrgomorpha cognata dominaient la plupart des sites.
Dans les milieux cultivés et les palmeraies, presque la moitié des espèces sont omniprésentes. Les milieux naturels se caractérisent par la catégorie d’espèces accessoires.
Les intervalles de classes de constance dans les neuf habitats décèlent six espèces rares : Calliptamus barbarus, Oedaleus senegalensis, Eunapiodes sp, Sphingonotus pachecoi, Phaneroptera nana et Conocephalus fuscus.
En général, la valeur de l’indice de la diversité de Shannon-Weaver, l’équitabilité et l’inverse de l’indice de Simpson évolue progressivement en Printemps et en Eté où elle est maximale, puis diminue en Hiver.
Cette diversité Alpha est élevée dans les sites cultivés moins perturbés et les palmeraies mal-entretenues. Par conséquent, les Orthoptères dans cette étude pourraient être utilisés avec succès comme bio indicateurs de la perturbation des habitats.
L’analyse factorielle des correspondances a permis de dégager trois tendances écologiques des Orthoptères. Les espèces qui préfèrent l’humidité vivent dans des zones moins ensoleillées et dans des parcelles recouvertes d’herbes denses telles que Phaneroptera nana, Conocephalus fuscus et Paratettix meridionalis.
Près de l’origine se trouve un groupe d’espèces relativement indifférentes aux facteurs d’humidité, d’ensoleillement et de couverture végétale, c’est le cas de Pyrgomorpha cognata et Pyrgomorpha conica.
Des espèces xérophiles qui préfèrent vivre dans des endroits secs, végétation très dispersée et un rayonnement solaire très important. Parmi ces espèces on cite entre autres : Sphingonotus rubescens et Sphingonotus pachecoi.
Le dendrogramme de similarité a divisé les habitats en 2 groupes : Un 1er groupe est formé par les milieux naturels (lits d’oueds) caractérisés par un faible taux de recouvrement végétal et sol rocailleux avec quelques endroits sablonneux, le 2ème groupe est formé par l’Acridofaune des milieux cultivés et palmeraies avec un recouvrement végétal assez important par apport aux milieux naturels, une irrigation plus au moins fréquente et le sol est sablonneux-limoneux.
L’évolution mensuelle des densités larvaires et d’adultes des populations acridiennes de la région d’étude a permis de décrire les cycles biologiques de 26 Caelifères. Nous avons pu déterminer des espèces à une seule génération, dont quatre espèces printanières et estivales et deux de la période Eté-Automne.
Les Acridiens à deux générations annuelles sont en nombre de onze, dont trois espèces reproduction continue, six à hivernation larvaire et deux espèces à hivernation embryonnaire.
Les Caelifères à trois générations annuelles sont représentés par cinq espèces. D’une manière générale on n’est pas parvenu à situer avec précision les différents phénomènes (éclosion, accouplement, ponte). Cela est dû, peut être à la méthode d’échantillonnage utilisée et surtout à la fréquence des relevés.
A la lumière de l’étude qualitative et quantitative du régime alimentaire des sept Acridiens, on distingue deux groupes de criquets : (i) Polyphages pour la majorité des espèces, dont Acrotylus patruelis, Heteracris annulosa, Aiolopus strepens, Pyrgomorpha cognata et Morphacris fasciata.
Ces espèces montrent un large spectre au niveau des plantes consommées, elles s’attaquent à plusieurs familles végétales toutes en marquants une tendance flagrante pour les Poaceae et en particulier Cynodon dactylon ; à l’exception de Heteracris annulosa et Pyrgomorpha cognata. (ii) Oligophages pour Acrida turrita et Ochrilidia gracilis.
Ces espèces ont consommées une seule famille végétale, les Poaceae. A travers le calcul du taux de consommation et l’indice d’attraction, on a constaté que les plantes les plus ingérées ne sont pas forcément les plus appétissantes. De plus, un végétal à faible recouvrement dans le biotope peut être attiré par le criquet.
Perspectives de recherche
Sur le plan taxonomique, il reste encore énormément d’aspects et de biotopes qui n’ont pas été exploitées et explorées, c’est pourquoi nous suggérons d’élargir ces recherches à d’autres milieux.
Il serait nécessaire d’approfondir les travaux systématiques en utilisant de nouvelles techniques comme la biologie moléculaire pour déterminer les espèces et sous- espèces.
La présente étude apporte son soutien à la suggestion que les sauterelles sont des indicateurs potentiellement utiles de perturbation des terres agricoles. De ce fait, il serait intéressant d’étudier les facteurs influençant la réponse des espèces Acridiennes à l’intensité des pratiques agricoles et l’abandon des parcelles.
Des mesures appropriées devraient être prises pour minimiser les risques de perturbation et de perte d’habitats naturels, car cela aurait des effets négatifs sur la composition et l’abondance de la communauté d’acridiens.
En effet, ce travail indique que les mauvaises herbes telles que Cynodon dactylon et Setaria verticillata abritaient à l’intérieur et autour des champs un grand nombre d’espèces de criquets.
Nous suggérons à ce que les plantes hôtes préférables par les Acridiens, soient cultivées autour du champ. Cela permettra de réduire les dommages aux cultures et de préserver la biodiversité des Orthoptères.
Pour une étude de phénologie des Acridiens plus détaillé on doit faire des relevés hebdomadaires pour ne pas échapper aux accouplements, aux pontes et aux éclosions des œufs.
Ces phénomènes vont déterminer avec précision le cycle biologique de chaque espèce d’Orthoptère. Pour réaliser un tel travail on recommande de s’intéresser uniquement à la biologie des Acridiens pour une future recherche.
En fin on peut conclure que toute amélioration de la prévision, donc de la lutte contre ces sautériaux, passe par une amélioration des connaissances des cycles biologique de ces populations.
Il est recommandé de mener des enquêtes similaires à grande échelle dans d’autres régions afin d’évaluer pleinement la faune d’acridiens de la région du Sahara septentrionale.
Il est grand temps de faire fait progresser le savoir sur les Acridiens à fin d’établir une liste nationale des Orthoptères sur laquelle l’on pourrait s’appuyer pour la gestion patrimoniale de la nature.