Chapitre III matériels et méthodes de travail
III – 1 – Matériel
III– 1 – 1 – Matériel utilisé sur le terrain
Pour la capture des Orthoptères nous avons utilisé un filet fauchoir. Il comprend un manche solide en roseau d’un mètre et demi de longueur portant sur l’une de ses extrémités un cercle métallique de 0,4m de diamètre. Un sac en toile est placé sur ce cercle métallique. Il est d’une profondeur de 0,5m (Figure 6b). Pour l’échantillonnage des criquets, nous avons délimité à l’aide d’une ficelle, des carrés de 3m de coté (Figure 6a). Les Caelifères récoltés sont placés dans des boites de Pétri chaque boite porte nécessairement une étiquette indiquant le lieu, la date et le numéro de la station. Les échantillons sont ramenés au laboratoire pour la détermination. Durant les sorties nous avons utilisé un carnet de prospection, dont lequel tous les renseignements concernant la sortie à savoir la date, le lieu, l’altitude, la pente, la température et la végétation, ainsi que toutes sortes d’informations sur le comportement des insectes sont notés.
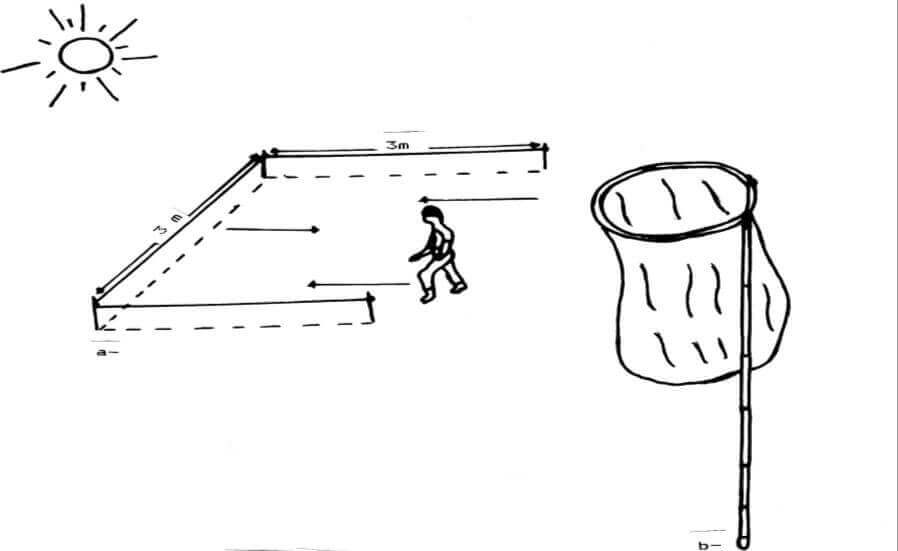
Figure 6. Méthode des quadrats : a)- carré d’échantillonnage ; b)- filet fauchoir
III – 1 – 2 – Matériel employé au laboratoire
III – 1 – 2 – 1 – Matériel utilisé pour la détermination des Orthoptères
Pour la détermination des espèces Orthoptéroloqiques nous avons utilisé la clé dichotomique de Chopard (1943) et les boites de collection des Orthoptères de l’insectarium du département de zoologie agricole de l’Institut National Agronomique d’El Harrach. Une loupe binoculaire est utilisée pour examiner avec précision les espèces acridiennes.
III – 1 – 2 – 2 – Matériel utilisé pour l’étude du régime alimentaire
Pour l’analyse des fèces nous avons utilisé 5 verres de montre. Ces derniers servent à contenir les différents produits pour l’étude du régime alimentaire. Des pinces pointes fines et lisses sont nécessaires à la trituration des fèces. Nous avons employé différents liquides de l’éthanol absolu de l’eau de javel, de l’eau distillée et du liquide de Faure. Les montages des épidermes sont réalisés entre lame et lamelle. Une plaque chauffante est nécessaire pour éliminer les bulles d’air existant entre lame et lamelle. Enfin pour pouvoir observer les différentes cellules végétales on doit utiliser un microscope photonique.
III – 2 – Méthode de travail
III – 2 – 1 – Méthodes utilisées sur le terrain
III – 2 – 1 – 1 – Choix des stations d’étude
Il convient de choisir la station ou site de prospection dans un biotope homogène suffisamment vaste de façon à éviter les effets de bordure et les interférences avec les biotopes voisins. Le site choisi doit être représentatif d’une catégorie de biotope largement représenté dans la région. Par ailleurs, il est impératif de suivre simultanément l’évolution des populations acridiennes dans plusieurs biotopes écologiquement contrastés en vue de faire des comparaisons. En effet, les peuplements acridiens seront différents et certaines espèces ne pourront être suivies que dans certains milieux car trop peu abondantes ou absentes dans les autres. L’humidité constitue, ainsi que nous l’avons vu, le principal facteur discriminant de la distribution des acridiens dans cette région. C’est pour cela nous avons choisi 3 stations dans des milieux d’humidités différentes, milieu non cultivé, milieu cultivé et une palmeraie. Pour représenter la physionomie de la végétation, nous avons jugé utile d’établir des transects végétaux pour chaque type de station. Nous avons délimité une aire d’échantillonnage de 10m sur 50m soit une surface de 500m2
La parcelle-échantillon de 500m2 est représentée suivant une projection orthogonale et en vue de profil. La projection verticale fournit des renseignements sur l’occupation du sol et sur la structure de la végétation. Ces notions sont complétées grâce à la représentation de profil qui donne une idée sur la physionomie du paysage. Celui-ci est de ce fait soit ouvert, semi fermé ou fermé. La connaissance de la physionomie du paysage est importante sachant que les Orthoptères qui sont assez héliophiles ne peuvent vivre en milieu fermé. Le taux de recouvrement des espèces végétales sur le terrain est estimé selon la méthode donnée par Duranton et al., (1982), qui consiste à estimer la surface de chaque espèce végétale en calculant la surface occupée par la projection orthogonale du végétal. La surface est déterminée grâce à la formule suivant :
π (d /2)2 x N T =
S
x100
T est taux de recouvrement d’une espèce végétale donnée
d est le diamètre moyen de la plante en projection orthogonale S est la surface du transect végétal soit 500m2
N est le nombre de touffes d’une espèce végétale donnée
Tableau 8 : Taux de recouvrement des espèces végétales dans le milieu non cultivé pour un transect de 500m2
| Espèces | Nombre de touffes | Diamètre en mm | Taux de recouvrement en % |
| Aristida pungens Aristida obtusa Colocynthis vulgaris Pituranthos chloranthus Pergularia tomentosa
Peganum harmala Arthrophytum scoparium |
3
79 2 7 4 2 9 |
1,10
0,15 0,75 0,60 0,85 0,55 0,80 |
0,57
0,30 0,20 0,40 0,45 0,09 0,14 |
| Total | 2,15 |
Tableau 9 : Taux de recouvrement des espèces végétales dans le milieu cultivé pour un transect de 500m2
| Espèces | Nombre de touffes | Diamètre en mm | Taux de recouvrement en % |
| Vitis vinifera Citrus sinensis Olea europea Capsicum annuum Mentha pulegium
Lycopersicum esculentum Lageneria vulgaris Cynodon dactylon Cyperus rotundus Setaria verticillata Echinops spinosus Aristida obtusa Amaranthus hybridus Tribulus terrester |
12
4 9 16 450 15 12 275 25 110 7 11 20 15 |
1,10
1,20 1,60 0,30 0,30 0,35 0,45 0,20 0,25 0,25 0,35 0,15 0,20 0,25 |
2,30
0,10 3,60 0,20 6,35 0,30 0,40 1,20 0,20 1,01 0,10 0,04 0,12 0,14 |
| Total | 17,06 |
Tableau 10 : Taux de recouvrement des espèces végétales dans la palmeraie pour un transect de 500 m2
| Espèces | Nombre de touffes | Diamètre en mm | Taux de recouvrement en % |
| Phoenix dactylifera | 8 | 1,60 | 3,20 |
| Setaria verticillata | 460 | 0,25 | 4,50 |
| Cynodon dactylon | 195 | 0,20 | 1,20 |
| Convolvulus arvensis | 55 | 0,35 | 1,05 |
| Peganum harmala | 4 | 0,60 | 0,20 |
| Total | 10,15 |
III – 2 – 1 – 2 – Transect végétal en milieu non cultivé
Le milieu non cultivé est situé à environ 5Km de Béni Isguen. C’est un reg encombré de cailloux, de sables grossiers et de limons plus fins. La végétation très éparse, est constituée principalement par une Chenopodiaceae arbustive à rameaux articulés. Il s’agit d’Arthrophytum scoparium. Le milieu non cultivé est caractérisé par son altitude, son exposition et sa pente :
Altitude : 530 m
Exposition : Sud
Pente : 0 %


Figure 7. Transect végétal en milieu non cultivé
Le transect végétal doit sa structure à une Chenopodiaceae arbustive précédemment citée. Le calcul du taux de recouvrement végétal donne une valeur faible de l’ordre de 2,157. (Fig.7, Tableau 8).
III – 2 – 1 – 3 – Transect végétal en milieu cultivé
Le milieu cultivé est situé à environ 7Km de Béni Isguen. C’est un terrain qui est mis en valeur. Les cultures sont installées sur des sols sablo-limoneux. Il y a comme cultures des arbres fruitiers tels que la vigne. L’oranger, le citronnier et l’olivier. Les Cultures maraichères sont représentées principalement par le piment, la courge, l’aubergine et la tomate. On y retrouve quelques plantes adventices comme le chiendent pied-de-poule. Cette station a les mêmes caractéristiques d’altitude, d’exposition et de pente que la station précédente
Altitude : 530 m
Exposition : Sud
Pente : 0 %

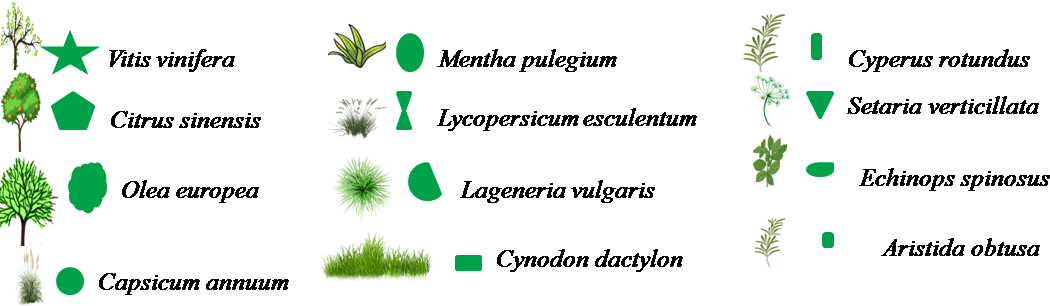
Figure 8. Transect végétal en milieu cultivé
Le transect végétal donne un profil à deux niveaux, un niveau occupé par les arbres et un autre par les plants de tomate, piment, courge et par les plantes adventices (Figure 8). La strate arbustive a une hauteur variant entre 1,5m et 2m. La strate herbacée quant à elle a une hauteur variant entre 0,1 et 0,5m. Le taux de recouvrement est estimé à 17,1 %, dû surtout à la menthe et à l’olivier qui présentent des valeurs respectives de 6,4% et 3,6% (Tableau 9). Pour les plantes adventices le taux de recouvrement le plus élevé est enregistré pour le chiendent-pied de poule avec une valeur égale à 1,3%.
III – 2 – 1 – 4 – Transect végétal dans une palmeraie
La palmeraie est située 5Km de Ghardaïa. La culture présente est le palmier dattier. Cette dernière est installée sur un sol sablo-limoneux. Au voisinage du palmier on trouve des plantes adventices dont la mieux représentée est la setaire verticillée, Setaria verticillata. La palmeraie d’étude diffère des deux stations précédentes par son exposition
Altitude : 530
Exposition : Nord
Pente : 0 %
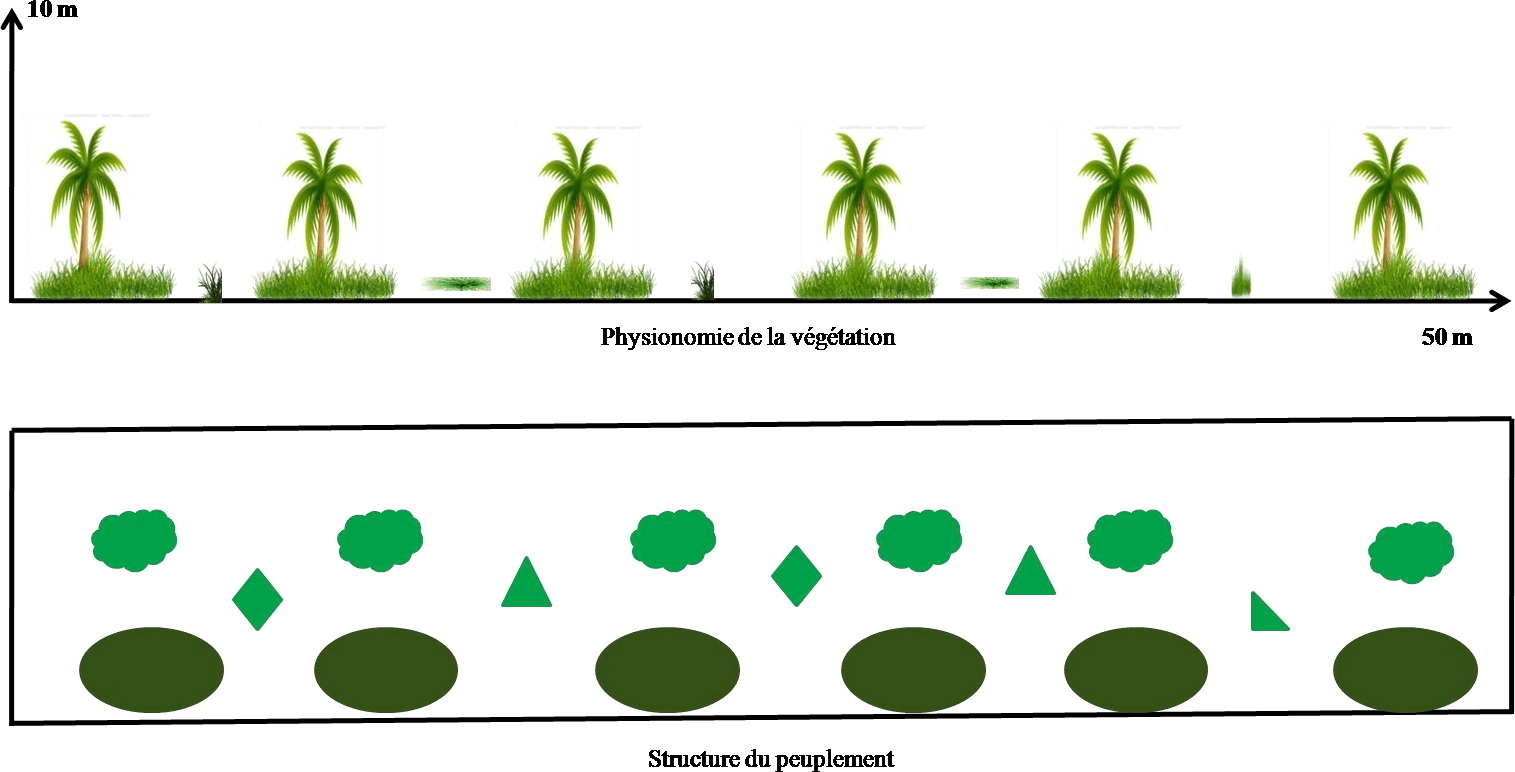

Figure 9. Transect végétal en palmeraie
Le transect végétal montre que le terrain est occupé uniquement par le palmier dattier et les mauvaises herbes (Figure 9). Le taux de recouvrement global de la station est égal à 10,2 % dont 4,5 % est celui de Setaria verticillata et 3,2 % du palmier dattier (Tableau 10).
III – 2 – 1 – 5 – Méthode des quadrats
Le but de l’échantillonnage est d’obtenir à partir d’une surface donnée, aussi restreinte que possible, une image fidèle de l’ensemble du peuplement (Lamotte et al, 1969). Plusieurs méthodes sont utilisées pour le dénombrement des populations d’Orthoptères. La méthode des quadrats est la plus pratique et qui donne des données exploitables. Selon Chessel et al., (1975) et Barbault (1981) le principe de cette méthode consiste à compter le nombre d’individus présents sur une surface déterminée pour obtenir une estimation satisfaisante de la diversité de la population. La surface d’échantillonnage dans laquelle nous intervenons est estimée à un hectare par station. Cinq prélèvements sont réalisés à l’intérieur de chaque station, dans des carrés de 9m2 chacun pris au hasard. Les prélèvements selon Voisin (1986) permettent de connaître la composition spécifique d’un peuplement d’Orthoptères.
III – 2 – 1 – 6 – Méthode d’étude du régime alimentaire sur le terrain III – 2 – 1 – 6 – 1 – Caractérisation de la végétation
Pour l’étude du régime alimentaire d’Acrotylus patruelis nous avons choisi le milieu cultivé décrit plus haut. Le taux de recouvrement des espèces végétales présentes dans un transect de 500m2 est calculé pour montrer l’importance de chaque espèce végétale (Tableau 9). De plus nous avons estimé l’abondance-dominance des espèces végétales présentes (Tableau 11). D’après Guinochet (1973), l’abondance-dominance est une expression de l’espace relatif occupé par l’ensemble des individus de chaque espèce, espace qui est déterminé à la fois par leur nombre et leurs dimensions. L’échelle la plus couramment utilisée est celle de Braun-Blanquet:
+ Simplement présent ou recouvrement très faible
1 Recouvrement faible de O à 5 % 2 Recouvrement de 5 à 25 %
Recouvrement de 25 à 50 %
Recouvrement de 50 à 75 %
Recouvrement de 75 à 100 %
Tableau 11. Abondance dominance des espèces végétales présentes dans le milieu cultivé
| Espèces végétales | Abondance-dominance | Espèces végétales | Abondances
dominance |
| Vitis vinifera Citrus sinensis Olea europea Capsicum annuum
Lycopersicum esculentum Lageneria vulgaris Cynodon dactylon Setaria verticillata Aristida obtusa Cyperus rotundus |
1
1 1 1 1 1 1 1 + 1 |
Mentha pulegium Echinops spinosus Sonchus oleraceus Erigeron canadensis Launaea resedifolia Salsola vermiculata Chenopodium album Amaranthus hybridus Portulaca oleracea Tribulus terrester | 2
+ + + + + + + + + |
III – 2 – 1 – 6 – 2 – Prélèvement des fèces
Les prélèvements des fèces ont lieu dans le milieu cultivé pendant deux années. Durant les mois allant de juillet à octobre de l’année 1992 nous avons effectué les premiers prélèvements. D’autres prélèvements ont été réalisés durant les mois de Juillet à septembre au cours de l’année suivante. Les criquets sont capturés entre 12 et 13 heures. Nous avons placé chaque insecte dans une boîte de Pétri. La durée suffisante pour que les acridiens vident leur tube digestif est variable selon les auteurs. Ben Halima et al., (1984), notent qu’il faut 7 heures pour récupérer les fèces après le repas d’un insecte. Par contre Launois (1976), signale que l’insecte doit jeûner 1 à 2 heures. Au contraire nous avons remarqué qu’il faut 24 heures pour vider le tube digestif d’Acrotylus patruelis. Les fèces de chaque individu sont conservées dans des cornets en papier, sur lesquels on inscrit le nom de l’espèce d’Orthoptère, le sexe de l’individu, la date et le lieu de capture.
III – 2 – 2 – Méthodes employées au laboratoire
III – 2 – 2 – 1 – Détermination des espèces capturées
Les échantillons ramenés du terrain font l’objet d’une détermination spécifique par le Professeur Doumandji à l’aide des clefs dont notamment celle des Orthoptères de l’Afrique du Nord de Chopard (1943). Voisin (1980), signale que contrairement aux imagos, les larves sont les plus souvent difficiles à identifier spécifiquement, même s’il existe de bons tableaux de détermination pour certains groupes.
III – 2 – 2 – 2 – Détermination des stades larvaires
La détermination des stades larvaires des différentes espèces est nécessaire pour l’étude de la dynamique de la population. Le nombre de stades larvaires n’est pas généralement facile à déterminer. Selon Duranton et al., (1982), le nombre réel de stades larvaires varie de 4 à 8 suivant les espèces. Le nombre le plus fréquent est de 5 à 6. Les 5 stades larvaires de la lignée mâle sont classés selon les étapes de développement des ébauches alaires : La larve de premier stade est caractérisée par une petite taille celle de l’œuf et par l’absence d’ébauches alaires. Celle de deuxième stade a des ébauches alaires A1 et A2, séparées et ne présentant pas de nervures. Chez la larve du troisième stade les ébauches alaires sont présentes, ne se recouvrent pas et montrent à ce moment là des nervures. Par contre les ébauches alaires de première paire A1 des larves du quatrième stade L4 recouvrent tout ou partie des ébauches alaires de seconde paire A2. Elles atteignent le quart ou le tiers de la longueur de l’abdomen Chez la larve du cinquième stade les ébauches alaires atteignent la moitié de la longueur de l’abdomen. La lignée femelle compte un stade larvaire supplémentaire. Les caractères de la larve L6 correspondent à ceux de la larve L5 futur mâle.
III – 2 – 2 – 3 – Conservation des échantillons
Les échantillons d’Orthoptères qui sont destinés à la collection sont tués dans un flacon contenant du coton imbibé d’acétate d’éthyle. Puis on les place sur des étaloirs en les fixant avec des épingles entomologiques au niveau du thorax, les ailes A2 et les élytres A1 sont maintenus dans une position horizontale, le bord postérieur des élytres faisant 90° avec l’axe du corps. Les étaloirs sont placés dans l’étuve à 45°C pendant quelques jours pour dessécher les Orthoptères. Après cela, ils sont retirés et placés dans une boite de collection. Une collection de référence est constituée au cours du déroulement des prospections Son but est de conserver un ou plusieurs individus de chaque espèce capturée dans les stations étudiées, généralement un mâle et une femelle par espèce. Cette collection sert de référence pour toute la durée des études et permet de vérifier les déterminations ultérieures.
III – 2 – 2 – 4 – Etablissement du catalogue des végétaux de référence
Dans le but d’établir un catalogue de référence on peut distinguer principalement deux méthodes. Celles-ci consistent à récolter à préparer et à photographier les fragments d’épidermes présents dans les fèces d’un animal nourri exclusivement sur une espèce végétale (Launois,1976), ou bien à prélever directement les épidermes des différentes parties de la plante et à les photographier (Chapuis, 1979; Butet, 1985; Ben Halima,1983) Nous avons employé la deuxième méthode citée qui offre l’avantage d’être rapide et qui permet surtout de savoir à quelle partie de la plante correspond l’épiderme étudié. Selon Butet (1985) l’obtention des épidermes peut se faire selon deux principes la séparation chimique et la séparation physique des épidermes. La séparation chimique des épidermes consiste à plonger des fragments végétaux dans des liquides de macération qui permettent de décolorer et de séparer les épidermes des tissus, tel que l’acide lactique. La deuxième méthode consiste en une séparation mécanique des épidermes. Les épidermes sont détachés délicatement des tissus sous-jacents avec de fines pinces ou quand cela n’est pas possible en plaçant l’épiderme à étudier en contact avec une lame de verre et en éliminant l’autre épiderme et les tissus internes par grattage. L’épiderme va passer dans de l’eau de Javel pendant 15 secondes. On fait passer l’épiderme dans de l’eau distillée pendant 2 minutes. Enfin les fragments épidermiques subissent des bains dans l’éthanol à concentrations progressives (70°, 90° et 100°). Les fragments épidermiques sont alors mis entre lame et lamelle dans du liquide de Faure pour l’observation au microscope photonique au grossissement 125×10. La collection de référence doit être la plus complète possible, tant au point de vue espèces, qu’organes de la plante, tige, feuille et inflorescence Nous signalons que c’est cette méthode que nous avons utilisée pour l’étude du régime alimentaire.
III – 2 – 2 – 5 – Analyse des fèces
La reconnaissance des débris végétaux contenus dans les fèces est facilitée par le ramollissement de celles-ci dans l’eau pendant 24 heures. L’ensemble passe ensuite dans une série de bains, dans de l’eau de javel, de l’eau distillée et de l’éthanol à différentes concentrations 70°, 90° et 100° Les montages se font dans une goutte de liquide de Faure entre lame et lamelle et sont examinés au microscope photonique (Figure 10). Selon Chara et al., (1986) l’analyse des contenus de fèces présente l’avantage de ne pas sacrifier les animaux ce qui peut être un inconvénient lorsque l’étude de l’alimentation est associée à une étude démographique de population, ou qu’elle concerne une espèce rares.
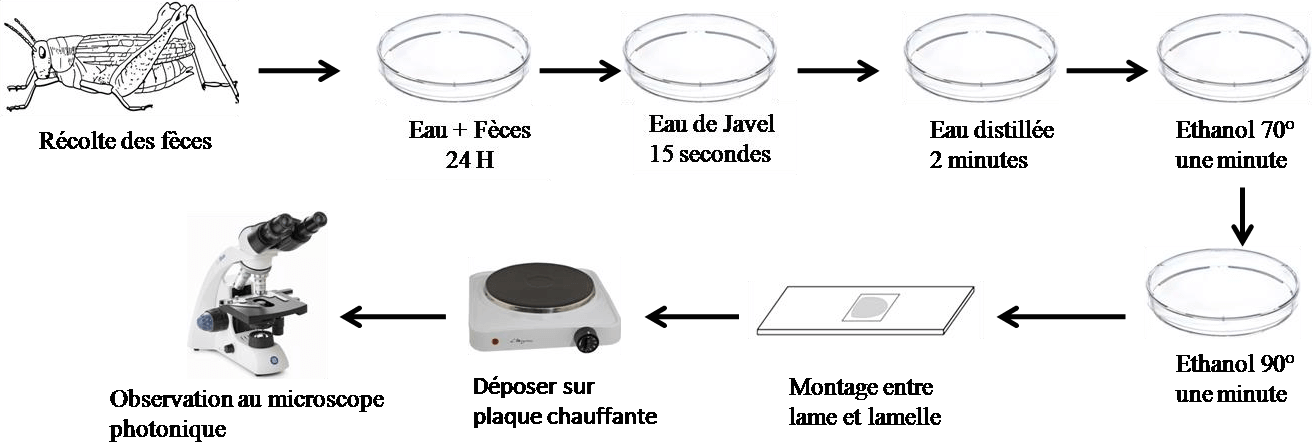
Figure 10. Démarche à suivre pour l’analyse des fèces
III – 2 – 3 – Méthodes d’expression des résultats
Les divers peuplements qui constituent une biocénose peuvent se définir quantitativement par un ensemble de descripteurs qui prennent en considération l’importance numérique des espèces qu’ils comportent (Ramade, 1984). Selon Voisin (1980), les individus d’une espèce donnée sont d’autant plus nombreux que les conditions écologiques auxquelles ils sont soumis, température, humidité et nourriture leur sont plus favorables.
III – 2 – 3 – 1 – Analyse statistique
III – 2 – 3 – 1 – 1 – Analyse factorielle des correspondances
C’est une méthode d’analyse multidimensionnelle qui permet d’établir un diagramme de dispersion unique dans lequel apparaissent à la fois chacun des caractères considérés et chacun des individus observés. Le résultat est obtenu grâce à une méthode particulière de codification et par un calcul de valeurs propres, qui assurent une parfaite symétrie entre les caractères et les individus, c’est à dire entre les lignes et les colonnes de la matrice des données initiales (Dagnelie, 1975). D’après Daget (1976), l’observation du graphique peut donner une idée sur l’interprétation des facteurs et montrer quelles variables sont responsables de la proximité entre telle ou telle observation.
III – 2 – 3 – 1 – 2 – Fréquence relative
C’est le pourcentage d’individus d’une espèce par rapport au total des individus. La fréquence relative peut être calculée pour un prélèvement ou pour l’ensemble des prélèvements d’une biocénose (Dajoz, 1971). Elle est représentée par La formule suivante :
Nombre d’individus de l’espèce
F =* 100
Nombre d’individus total
III – 2 – 3 – 1 – 3 – Constance
C’est le rapport exprimé sous forme de pourcentage (Dajoz, 1982) : C = P * 100 / R dans lequel, P est le nombre de relevés contenant l’espèce étudiée et R le nombre de relevés effectués. En fonction de C on distingue :
Les espèces constantes présentent dans plus de 50 % de relevés
Les espèces accessoires présentes dans 25 à 50 % de relevés
Les espèces accidentelles trouvées dans moins de 25 % de relevés.
III – 2 – 3 – 2 – Analyse par indices écologiques
III – 2 – 3 – 2 – 1 – Indice de diversité et équirépartition
De nombreux indices de diversité sont proposés, et selon Frontier (1982), l’indice le plus communément utilisé aujourd’hui est celui de Shannon-Weaver, dérivé de la théorie de l’information.
i = s
H’ = – ∑ pi* log2 pi i =1
Où S est le nombre d’espèces et pi représente la probabilité de rencontre de l’espèce de rang
i. On peut écrire pi = ni/N, où ni est l’effectif de chaque espèce dans l’échantillon et N la somme des ni toutes espèces confondues. H’ est l’indice de diversité de l’échantillon. Il est exprimé en unités d’information par individu, ou bits par individus. Le logarithme utilisé est de base 2. On démontre que H’ est bien maximal, pour un nombre d’espèces S donné quand toutes ces espèces sont également fréquentes. Cette valeur maximale est H’max = log2 S. Enfin la connaissance de H’ et H’max permet de déterminer l’équitabilité E.
H’H’
E == H’maxlog2 S
E varie entre 0 et 1. Quand E est inférieur à 0,5, ceci traduit un déséquilibre au sein d’un peuplement où une ou deux espèces pullulent par rapport aux autres. Si E est supérieur à 0,5 il s’établit un équilibre entre les différentes populations composant un peuplement. Selon Dajoz (1985), un indice de diversité élevé correspond à des conditions de milieu favorables permettant l’installation de nombreuses espèces. Une diversité élevée révèle alors une stabilité plus grande. Une équitabilité élevée est l’indice d’un peuplement équilibré.
III- 2- 3- 2- 2- Indice de dispersion des espèces acridiennes dans le temps et dans l’espace
L’étude de la répartition ou dispersion permet de déterminer le mode de distribution des Orthoptères dans les différentes stations en fonction du temps. Selon Bachelier (1978), si N est le nombre total d’observations, m la moyenne des comptages et X le résultat d’un comptage quelconque, la variance peut s’écrire :
∑ (X – m) S2 =-
N – 1
Si m > S2 on a une distribution régulière
Si m = S2 on a une distribution au hasard ou aléatoire
Si m < S2 on a une distribution en agrégats dite encore distribution contagieuse Si S2 = O la distribution est uniforme (Dajoz, 1971).
III – 2 – 3 – 3 – Méthodes de quantification relative de la nourriture ingérée
Butet (1985), a recensé diverses méthodes de quantification relative de la nourriture ingérée par des phytophages; elles sont basées sur le dénombrement ou la mesure des surfaces des fragments épidermiques présents sur les lames échantillons. Nous pouvons distinguer les principaux cas suivants :
Tous les fragments présents sur la lame-échantillon sont dénombrés (Launois, 1976)
Un nombre prédéterminé de fragments est recensé par un balayage méthodique continu de la lame-échantillon (Chapuis, 1979).
Les surfaces d’un nombre prédéterminée de fragments sont mesurées (Nel et al, 1973 in Butet, 1985).
Le dénombrement ou la mesure des surfaces des épidermes sont effectués dans des zones définies sur la lame grille d’observation espace entre deux lignes, champs de microscope, etc.…
Pour l’expression des résultats du régime alimentaire d’Acrotylus patruelis nous avons utilisé deux méthodes.
III – 2 – 3 – 3 – 1 – Méthode des fréquences
Butet (1985), définit une fréquence relative (F %) d’apparition d’un item donné dans les échantillons :
ni
F % item i = —— x 100
N
Où ni est le nombre d’échantillons où l’item i est présent et N est le nombre total d’échantillons pris en compte.
III– 2 – 3 – 3 – 2 – Méthode des surfaces
Le principe de la méthode consiste à calculer la surface ingérée en millimètres carrés pour chaque espèce végétale. Pour cela nous avons utilisé un carré, Fenêtre d’un millimètre carré. Le papier millimétré est collé sur le plateau du microscope photonique, de façon à ce que l’objectif soit en face du carré. On place le montage des fèces sur le papier millimétré, puis on procède à un balayage de toute la surface de la lamelle. On note la surface des fragments végétaux qui occupent chaque carré. Pour montrer l’aspect quantitatif des espèces végétales ingérées nous avons utilisé les formules suivantes proposées par Doumandji et al., (1993).
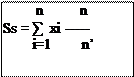
![]()
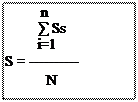
![]()
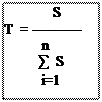
![]()
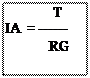
Ss : est la surface d’une espèce végétale donnée rejetée dans les fèces et calculée pour un individu.
Xi : est la surface des fragments du végétal de l’espèce i notée dans les fèces d’un individu. n : est le nombre de mm2 de la lamelle soit 576 mm2.
n’ : est le nombre de mm2 observés sur la lamelle vides ou occupés par les fragments végétaux. Le rapport n/n’ délimite le champ de travail et permet de diminuer les erreurs de manipulation.
S : est la surface moyenne d’une espèce végétale consommée par N individus.
n
∑ S : est la somme des surfaces moyennes des végétaux rejetées par individu toutes i=1végétales confondues.
N : est le nombre d’individus pris en considération.
T : est le taux de consommation pour une espèce végétale par rapport à l’ensemble des surfaces végétales rejetées.
IA : est l’indice d’attraction d’une espèce végétale donnée.
RG : est le recouvrement global pour une espèce végétale présente dans la station d’étude.