Le cycle biologique des acridiens (orthoptères)
I-1-4-Cycle biologique
Les acridiens passent toujours par trois états biologiques au cours de leur vie, les états d’œuf, de larve et d’imago (Fig.1).
Le terme adulte est réservé aux individus physiologiquement capables de se reproduire (Appert et Deuse, 1982).
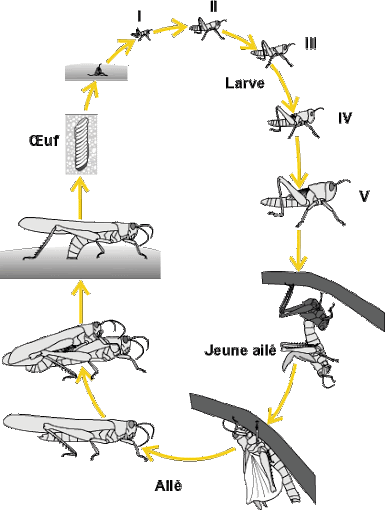
Figure 1. Cycle biologique du criquet pèlerin Schistocerca gregaria Forskal, 1775 (COPR, 1982)
I-1-4-1-Œufs
Dans les régions tempérées la plupart des Orthoptères pondent pendant la belle saison et l’éclosion des oeufs se produit après une incubation très variable, allant de quelques Jours à plusieurs mois et pouvant même dépasser une année chez certaines espèces (Chopard, 1943).
I-1-4-1 – 1-Ponte
La ponte des Orthoptères se fait le plus souvent dans la terre, bien que quelques très rares espèces déposent leurs oeufs dans les tissus végétaux.
La femelle, après avoir rencontré l’endroit qui lui convient, se dresse très haut sur ses quatre pattes antérieures et médianes, l’abdomen fortement arqué, l’extrémité étant perpendiculaire à la surface du sol.
L’action de l’oviscapte chez les Caelifères est très différente de celle des Ensifères (Chopard, 1949).
C’est par des mouvements alternatifs des valves de l’oviscapte que la femelle arrive à forer un trou de 6 à 10cm de profondeur où elle enfonce son abdomen.
Celui-ci s’allonge jusqu’à égaler 2 à 3 fois la longueur initiale de son corps. Chambille (1977), a étudié le comportement de forage des femelles du Criquet migrateur africain en milieu humide et sec.
Il a remarqué qu’en milieu humide la ponte s’effectue après peu d’essais infructueux. Les forages sont superficiels et n’atteignent pas 3cm de profondeur, distance correspondant à la pénétration de l’abdomen dans le substrat.
Lorsque le forage est profond supérieur ou égal à 3cm, il est presque toujours suivi du dépôt de l’oothèque. En milieu sec, les essais sont nombreux pour toutes les pontes.
La profondeur moyenne du forage est significativement plus grande. Elle résulte d’une augmentation non négligeable des essais infructueux profonds supérieur ou égal à 3cm. Les oeufs sont pondus soit isolément soit en masse.
Dans le deuxième cas la femelle commence par déposer un mucus de mucco-polysaccharides au fond du trou. Puis elle pond ses oeufs qui sont agglomérés par le mucus.
La ponte se termine par un bouchon spumeux ayant la même origine. Le nombre d’oeufs présents dans une oothèque varie suivant les espèces allant de 10 à 100 oeufs.
Parfois la femelle peut déposer plus d’une oothèque, 2 ou 3 en général, parfois une dizaine au cours de sa vie (Chopard, 1938).
I-1-4-1 – 2-Incubation
La durée de l’incubation ou le développement embryonnaire est variable selon les espèces et les conditions climatiques. Selon Nurein (1989), les œufs de Schistocerca gregaria ont besoin d’une humidité suffisante pour se développer.
Ils puisent de l’humidité dans le sol. Les larves de cet Orthoptère ne peuvent donc éclore qu’à partir des œufs déposés dans le sol. Si l’humidité nécessaire au développement des oeufs fait défauts ceux-ci peuvent survivre jusqu’à 60 jours.
La période d’incubation des oeufs dépend aussi de la température du sol.
L’incubation dans les champs dure environ 30 jours. Lorsque les températures sont élevées, elle ne dure que 23 jours alors que pendant la saison froide, avec des températures basses, elle peut s’étaler jusqu’ à 60 jours.
I-1-4-2-Développement larvaire
Les larves vivent à la surface du sol, dans les herbes, les arbustes et rarement dans les arbres (Appert et Deuse, 1982).
I-1-4-2 – 1-Eclosion
Dans le cas le plus général, au moment de l’éclosion, l’oothèque étant enfouie dans le sol les larves néonates cheminent à travers le bouchon spumeux et sortent à la surface du sol à l’état de larves rampantes.
Elles se déplacent quelques minutes seulement comme un ver; ce sont les larves du stade rampant; chaque larve est encore emmaillotée dans la membrane interne de l’œuf ou aminios ; elle s’en dégage rapidement; c’est la fausse mue (Chopard, 1938).
I-1-4-2 – 2-Nombre de stades larvaires
En général il y a cinq stades larvaires mais ce nombre peut varier en fonction des espèces et du sexe.
Pour le Criquet nomade Nomadacris septemfasciata (Serville, 1838) Ndyanabo et Byaruhanga (1969), signalent 6 ou 7 stades larvaires.
La période larvaire coïncide avec la croissance active de la végétation qui procure aux criquets la nourriture et la protection nécessaires. La phase solitaire comprend 7 stades larvaires tandis que la phase grégaire n’en compte que 6. Le développement complet, depuis l’éclosion de l’oeuf jusqu’à l’insecte ailé dure 2 mois à 2 mois et demi dans des conditions favorables.
Les larves de Criquet pèlerin passent, de l’éclosion à l’état imaginal, par plusieurs stades larvaires. Leur nombre est variable en fonction de la phase: 5 stades chez les grégaires et 6 le plus fréquemment chez les solitaires.
Le stade supplémentaire se situe entre le troisième et le quatrième stade (Duranton et Lecoq, 1990).
Selon Duranton et al. (1982) le nombre de stades larvaires n’est pas toujours connu avec certitude sauf pour les ravageurs les plus importants et les plus étudiés comme Nomadacris septemfasciata, Locusta migratoria, Schistocerca gregaria et certains acridiens qui présentent la particularité d’avoir un nombre de stries oculaires proportionnel au nombre de stades larvaires.
La durée du développement larvaire varie essentiellement en fonction de la température de l’air.
Par ailleurs, dans des conditions écologiques identiques les grégaires se développent plus rapidement que les solitaires.
Chez les grégaires dans de bonnes conditions, la durée de développement larvaire la plus courte est de 25 jours. Dans de mauvaises conditions elle peut atteindre 50 jours.
Chez les solitaires, dans des conditions optimales, le développement larvaire dure au minimum 30 jours. Il peut s’étendre à trois mois en cas de conditions très défavorables (Duranton et Lecoq, 1990).
I-1-4-3-Imago
La dernière mue donne naissance à un imago. La première partie de la vie imaginale est surtout consacrée à la recherche d’un biotope favorable et à l’alimentation.
Mâles et femelles augmentent de poids dans des proportions notables accumulant du corps gras.
Puis le poids des mâles se stabilise, alors que celui des femelles continue à augmenter. Ce deuxième accroissement du poids est en rapport avec la maturation ovocytaire préparant la future première ponte.
Lorsque les ailés sont en période de reproduction, on parle d’adultes (Duranton et al., 1982).
Pasquier (1946) a présenté les différents âges de Schistocerca gregaria à l’état imaginal. Selon l’auteur précité, on doit distinguer d’abord les ailés immatures des adultes vrais génétiquement parfaits, le mot adulte étant alors pris dans le sens strict d’insecte en état de reproduction.
Par analogie à une nomenclature utilisée par les ichtyologues les derniers seront appelés génétiques sachant que le terme agénétique désigne les immatures.
Dans l’Afrique du Nord, en été, à la suite de la mue imaginale, les ailés roses Jeunes ou népiogones se trouvent réalisés. Aux népiogones, succède un âge de durée et d’importance économique variables, celui des errants roses ou néogones.
Quoiqu’il soit, au bout de peu de temps, les néogones, en vol élevé, profitant certainement de courants aériens favorablement orientés, quittent l’Afrique du Nord pour effectuer la traversée assez rapide du Sahara.
En quelques jours ces hypogénétiques ou migrants roses, dits de retour (émigrants pour les Nord-africains) gagnent des territoires méridionaux dont certains sont propices à leur maturation sexuelle. Devenant génétiques (éogénétiques), ils jaunissent sans rougir. Les ailés effectuant leur premier accouplement et leur première ponte sont appelés néogénétiques.
Par la suite, ils seront désignés par eugénétiques pendant toute la période de reproduction.
Vers la fin de leur vie, en juillet, les ailés vieillie sont gérogénétique, les males étant jaunes paille, les femelles plus brunes à abdomen ardoisé ou violacé.
I-1-4-4-Accouplement
L’époque laquelle l’accouplement a lieu est variable, suivant les espèces et elle est naturellement liée au moment où les insectes deviennent adultes, c’est à dire sexuellement mûrs (Chopard, 1938).
Le rapprochement des sexes est préparé chez un certain nombre d’Orthoptères par des manifestations liées à la période d’excitation sexuelle.
La plus remarquable de ces manifestations est la stridulation (Chopard, 1943).
En général, pendant l’accouplement le mâle se trouve sur la femelle, de telle façon que ses pattes antérieures s’accrochent à l’angle antérieur des lobes latéraux du pronotum il abaisse et recourbe son abdomen; la plaque sous-génitale est écartée et le pénis se trouve dégagé de son prépuce avant d’être introduit entre les valves de l’oviscapte; les cerques du mâle jouent un rôle en pinçant la plaque sous-génitale de la femelle (Chopard, 1949).
I-1-4-5-Nombre de générations
Le nombre de générations annuelles qu’une espèce peut présenter correspond au voltinisme. On distingue des espèces univoltines, n’effectuant qu’une seule génération dans l’année et des espèces polyvoltines à plusieurs générations annuelles.
Parmi ces dernières, on sépare les espèces en bivoltines, en trivoltines et en tétravoltines selon qu’elles aient 2, 3 ou 4 générations par an.
Le nombre maximal de générations qu’une espèce peut effectuer en une année semble être de 5 chez les acridiens, encore que ces cas soient assez rares. A l’opposé, on connaît des espèces qui ont besoin de deux années au moins pour effectuer un cycle complet, particulièrement dans les régions froides et très arides.
En zone tropicale sèche, les acridiens présentent en majorité de 1 à 3 générations par an.
Pour une même espèce, le nombre de générations peut être variable suivant la région dans laquelle la population se développe ou en fonction des caractéristiques météorologiques annuelles (Duranton et al., 1982).
D’après Gillon (1989), plusieurs espèces dangereuses ne possèdent qu’une génération par an; elles passent la saison sèche à l’état d’oeuf ou d’ailé immature.
A l’opposé, chez les espèces les plus à craindre en Afrique comme le Criquet pèlerin et le Criquet migrateur, il peut y avoir quatre à cinq générations successives dans l’année si les conditions écologique s’y prêtent.
Profitant des crues saisonnières du Niger pour prolonger sa période annuelle de reproduction, le Criquet migrateur africain peut avoir par exemple Jusqu’à 4 ou 5 générations au Mali.
Dans les conditions normales de sécheresse, il n’y a que 2 générations.
En colonisant d’autres habitats dans certaines régions agricoles, comme c’est le cas notamment, en Afrique du Sud, ce Criquet est capable de développer 3 à 4 générations, ce qui le rend problématique (Mason, 1989).