
Les étapes d’implantation embryonnaire
II- Les étapes d’implantation embryonnaire
II-1- L’étape de la fécondation (implantation)
La rencontre entre le spermatozoïde et l’ovocyte est une des étapes essentielles de la reproduction. Elle nécessite un ovocyte et un spermatozoïde tous deux compétents.
Elle se déroule généralement au tiers moyen supérieur de la trompe de Fallope.
Lors de la fécondation, le spermatozoïde entre en contact avec la zone pellucide entourant l’ovocyte, cette zone pellucide sert de barrière empêchant la polyspermie.
La première division a lieu très rapidement (durant les 24 premières heures) et se déroule toujours dans la trompe. L’embryon n’atteint l’utérus que durant le 5ème jour de son développement.
Il reste entouré des cellules de la corona radiata jusqu’au stade blastocyste (Virginie Gridelet,2015).
II-2- Division cellulaire (segmentation)
Les facteurs maternels précoces jouent un rôle primordial durant le début du développement embryonnaire.
L’ovocyte supporte les premières divisions du zygote et fournit le matériel protéique nécessaire jusqu’au stade 4-8 cellules.
Il apporte la grande majorité du cytoplasme et des composants nucléaires de l’embryon.
Les cellules embryonnaires, aussi appelées blastomères, sont flexibles et totipotentes, autrement dit, elles sont capables en se développant et en se multipliant pour donner tous les types cellulaires du corps humain (De Paepe et al., 2014).

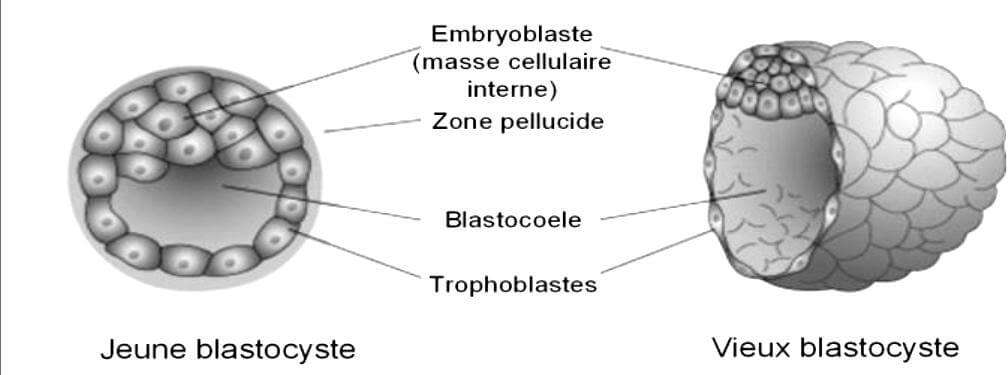
II-3 Le blastocyste (implantation)
Au stade blastocyste, l’embryon sort de la zone pellucide qui l’entoure et ses cellules s’organisent, pour la première fois, en deux types cellulaires différenciés:
- le premier, appelé trophoblaste, qui forme une couche unicellulaire et forme le pourtour du blastocyste,
- le second type forme la masse cellulaire interne cellules qui donnent le futur fœtus.
Le trophoblaste est à l’origine du chorion qui forme le placenta et la séparation entre l’embryon et les cellules maternelles (De Paepe et al., 2013).
II-4 Implantation embryonnaire
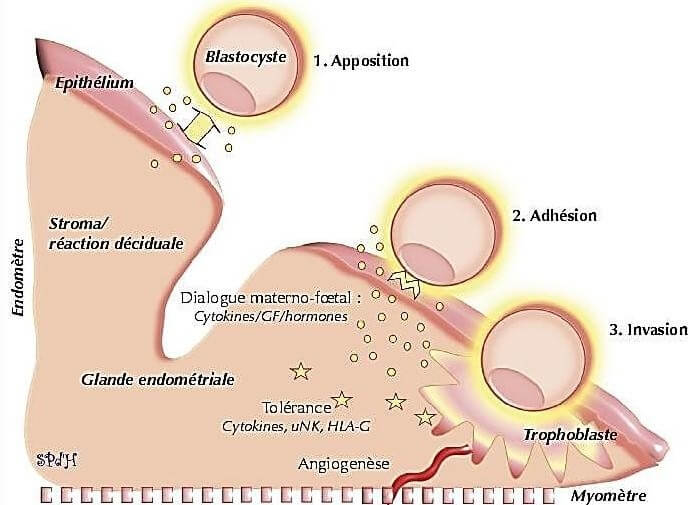
Le succès de l’implantation embryonnaire n’est pas uniquement une préparation hormonale adéquate de l’endomètre mais nécessite une interaction étroite, synchronisée et réciproque entre les différentes cellules de l’endomètre et l’embryon.
C’est ce qui est appelé dialogue materno-fœtal durant la phase d’implantation.
Ce dialogue s’établit grâce à de nombreuses cytokines, facteurs de croissance, chimiokines, récepteurs et molécules d’adhésion produits et sécrétés par l’endomètre et par l’embryon (Thouas et al., 2014).
La plupart des grossesses biochimiques et les fausses couches spontanées, approximativement deux tiers, sont dues à une implantation incomplète de l’embryon (Norwitz et al., 2000).
L’éclosion embryonnaire permet un contact direct avec les cellules épithéliales de l’endomètre.
Ce contact, appelé phase d’apposition. Durant cette phase, de véritables contacts dynamiques ont lieu entre le blastocyste et l’endomètre via des molécules d’adhésion et des récepteurs :
- L’interleukine-1 (l’IL-1) : sécrété par le blastocyste augmente l’expression d’intégrines à la surface de l’endomètre lui permettant d’interagir avec l’embryon.
- La mucine MUC-1 : l’épithélium est recouvert sur toute sa surface épithéliale de glycocalyx, expliquant en partie sa nature non adhésive.
Au niveau du site d’implantation, une diminution de l’expression des mucines (principalement de MUC-1) à la surface des cellules épithéliales endométriales est observée et serait impliquée dans le positionnement du blastocyste. (Brayman et al., 2004).
- L’expression de selectines à la surface du blastocyste ainsi que celles de sélectines complémentaires à la surface de l’épithélium endométrial durant la phase réceptive (Genbacev et al., 2003).
Après la phase d’apposition vient la phase d’adhésion, durant laquelle l’embryon et l’endomètre ont des contacts plus précis via des molécules d’adhésion (et principalement des intégrines).
Le trophoblaste se sépare en deux types cellulaires distincts : le syncytiotrophoblaste et le cytotrophoblaste (Gridelet, 2015).
Le syncytiotrophoblaste forme la couche extérieure des futures annexes embryonnaires qui est en contact avec les cellules épithéliales de l’endomètre et la lumière de la cavité utérine.
Le cytotrophoblaste forme une couche interne qui est en contact avec la masse cellulaire interne et le blastocœle.
Le syncytiotrophoblaste sécrète des enzymes lytiques et des facteurs déclenchant l’apoptose des cellules épithéliales de l’endomètre pour permettre l’invasion de l’embryon au sein de l’endomètre qui correspond à la dernière phase de l’implantation (Gridelet, 2015).
L’invasion du blastocyste dans l’endomètre commence par la liaison des cellules du syncytiotrophoblaste aux constituants de la membrane basale et de la matrice extracellulaire de l’endomètre via les intégrines.
Le trophoblaste envahit ensuite le stroma conjonctif sous-épithélial suite à une décomposition des composants de la membrane basale et de la matrice extracellulaire, décomposition liée à l’activation de métalloprotéases.
L’endomètre joue donc un rôle important par l’expression de protéines d’ancrage et par sa modification en décidue lors de l’implantation.
La placentation débute après l’enfouissement total de l’embryon au sein de l’endomètre et le placenta se forme suite à l’apparition de villosités dans le trophoblaste (B. Gellersen, I. A. Brosens, J. J. Brosens,2007).
II-5 Molécules et cellules importantes dans le dialogue materno-fœtal
À côté des molécules d’adhésion qui représentent la « partie visible » du dialogue existant entre le blastocyste et l’épithélium endométrial, toute une série de molécules et de cellules sont impliquées dans la régulation de l’implantation.
II-5-1 L’hCG (hormone Chorionique Gonadotrope)
L’hormone chorionique gonadotrope (hCG) est une hormone glycoprotéique de 36 à 40 kDa. Elle est composée de deux sous-unités, α et β liées par une liaison non covalente.
La sous-unité α, composée de 92 acides aminés, est codée dans le chromosome 6 et est commune aux différentes hormones de la famille des glycoprotéines dont font partie la LH, la FSH et l’hormone thyréostimuline( TSH) (P. F. Policastro, S. Daniels-McQueen, G. Carle, I,1986).
La sous-unité β, qui est différente pour chaque hormone, est codée sur différents gènes situés sur le chromosome 19 (LH, hCG et TSH) ou sur le chromosome 11 (FSH).
La sous-unité β de l’hCG est codée dans 6 gènes différents mais très similaires situés dans un groupe de gènes sur le chromosome 19.
L’hCG a une structure similaire à la LH, mais contrairement à elle, l’hCG existe sous plusieurs formes, connues comme l’hCG classique, l’hCG hyper-glycosylée et l’unité β libre de l’hCG hyper-glycosylée (Cole, 2009 ; Fournier et al., 2015).
Chacune de ces 4 molécules ont des fonctions physiologiques différentes.
Cette hormone a été appelée hormone chorionique gonadotrope. La stimulation de la production de la progestérone par le corps jaune a été longtemps la seule fonction connue de l’hCG.
La glycosylation des hormones aurait une influence directe sur leur bioactivité. L’hCG hyper-glycosylée interviendrait d’avantage dans l’implantation que l’hCG classique (Cole, 2009).
Trois équipes indépendantes ont montré que le blastocyste pré-implantatoire sécrète lhormone Chorionique Gonadotrope dans l’espace utérin qui se lie à son récepteur hCGR sur la surface déciduale.
En réponse, la décidue se prépare pour l’implantation (Srisuparp et al., 2001 ; D’Hauterive et al., 2004).
L’hCG augmente la sécrétion du Leukemia Inhibitory Factor (LIF) et diminue celle de l’interleukine -6 par les cellules endométriales, molécules connues pour leur influence sur l’implantation embryonnaire (Perrier d’Hauterive et al., 2004).
Il favorise la différenciation des cytotrophoblastes en synciotiotrophoblastes. Il peut réguler la synthèse des prostaglandines et la formation de l’Adénosine mono-phosphate cyclique (AMPc).
Une étude récente a montré que les différentes formes d’hCG stimuleraient l’invasion trophoblastique indépendamment du récepteur classique à l’hCG, le LHCGR (Lee et al., 2013).
L’hormone Chorionique Gonadotrope a également des actions angiogéniques et immunologiques, il augmente la formation des vaisseaux sanguins et la migration et la maturation des péricytes (Berndt et al., 2006 ; Bourdiec et al., 2013).
L’hCG assure au placenta un approvisionnement en sang maternel adéquat durant son invasion de l’endomètre fonctionnel avec une nutrition optimale du fœtus. L’hCG régule la prolifération des cellules Natural killer utérines ( uNK) (Bansal et al., 2012).
Ces cellules n’expriment pas le LHCGR et l’hCG agirait directement sur ces cellules par un autre récepteur, le mannose récepteur qui est exprimé par les uNK (Kane et al., 2009) .
L’hormone Chorionique Gonadotrope hCG agit également sur d’autres cellules immunitaires, les monocytes en promouvant leur fonction et leur sécrétion d’IL-8 (Kosaka et al., 2002) ,et aussi induisant les fonctions des macrophages (Wan et al., 2007).
En stimulant la fonction des macrophages, l’hormone Chorionique Gonadotrope assainit l’endomètre en l’épurant des cellules apoptotiques et en le défendant d’éventuelles infections, deux mécanismes importants pour le maintien de la grossesse.
Par ailleurs, l’hCG influence la différentiation et la fonction des cellules dendritiques, diminuant leur capacité à stimuler la prolifération des lymphocytes T (Wan et al., 2008).
Au niveau des cellules nucléaires du sang périphériques, l’hCG induit la sécrétion de différentes cytokines, dont l’IL-1β et LIF et pourrait stimuler l’invasion trophoblastique (Yu et al., 2015).
Finalement, l’hCG a différents effets sur les lymphocytes T (Cluster de Différenciation CD4+) , avec un impact positif sur la prolifération des cellules T CD4+25+ et en attirant ces cellules vers l’endomètre en début de grossesse (Khil et al., 2007 ; A. Schumacher et al., 2009).
Des études récentes montrent par ailleurs qu’une trop grande quantité d’hormone Chorionique Gonadotrope, telle qu’utilisée en procréation médicalement assistée ( PMA ) pour déclencher l’ovulation, serait délétère pour la réceptivité utérine, tout est une question d’équilibre (Evans et Salamonsen, 2013).
II-5-2 La Hormone Lutéinisante (LH)
La LH est sécrétée par la glande pituitaire et est une glycoprotéine hétéro-dimèrique avec une masse moléculaire de 28kDa comprenant deux sous-unités.
Cette hormone a un rôle essentiel dans la reproduction en induisant la maturation ovocytaire et en provoquant l’ovulation durant le cycle menstruel.
La Hormone Lutéinisante LH contrôle la production de la progestérone par le corps jaune et si une grossesse survient chez l’être humain, l’hCG sécrétée par les cellules embryonnaires supplante le rôle de la LH pour la régulation de la production de la progestérone.
Pour l’établissement de la grossesse, la Hormone Lutéinisante LH favorise la décidualisation de l’endomètre.
Des scientifiques ont exploré les propriétés d’adhésion des lymphocytes dans l’utérus murin et ont révélé que cette adhésion était plus grande en présence de LH.
Ces auteurs ont suggéré que la LH pourrait activer les molécules d’adhésion à la surface des précurseurs des cellules uNK, ainsi qu’augmenter leur recrutement dans l’utérus.
Ils suggèrent encore que la LH pourrait avoir un impact sur le recrutement des cellules uNK (Van Den Heuvel et al., 2005)